
|
You have probably heard of at least one eight-legged femme fatale: the black widow. A female so fearsome she has inspired a multitude of book, movie and comic book characters. It turns out that this aggressive arachnid is not alone! Spiders are a group rich with outsized females. In biology, when one sex is differently sized than the other within the same species it is called sexual size dimorphism, or SSD. SSD can be male or female-biased, meaning either the male or female can be abnormally sized. Female-biased SSD, more common in egg-laying organisms, has evolved repeatedly in spiders. In fact, female-biased SSD has evolved four to nine times in the orb weaver group alone (Hormiga et al., 2000; Kuntner et al., 2015). Dr. Matjaž Kuntner, the Chair of the Biological Institute at the Research Centre of the Slovenian Academy of Sciences and Arts, studies the phenomenon of female-biased SSD in orb weaver spiders and has made several fascinating discoveries. He first became interested in SSD during his graduate fieldwork in the tropics while earning his doctorate at George Washington University. While studying orb weaver systematics in places such as Southeast Asia, Madagascar and South Africa, he witnessed the obvious size discrepancies between males and females. He fell into the UMD web when he took Dr. Jeff Shultz’s course, Arthropod Form and Function, in our own Department of Entomology.
Both behaviors indicate that Nephilids exhibit something called sexual conflict. This is when males and females have opposing goals and strategies during mating and can lead to the sexes evolving ever more complicated ways to outsmart and outmaneuver one another (Kuntner et al., 2015). While Dr. Kuntner’s lab works primarily on Nephilid behavior, the Argiopines also exhibit sexual cannibalism and emasculation, indicating sexual conflict could play an important role in this group as well (Nessler et al., 2009). One of Dr. Kuntner’s main goals is to understand the evolutionary patterns of female-biased SSD. In these systems, females are much larger than males, which begs the question: are females giants or are males dwarves? We assume that the ancestors of groups that exhibit SSD were once monomorphic, meaning males and females were the same size. Varying selection types and selection pressures lead to various iterations of SSD. Using analysis of evolutionary history, Dr. Kuntner was able to determine that the evolution of male and female body size is actually decoupled in Nephilids. Over time, females and males have both independently increased in size. However, females have undergone much greater increases over the same amount of evolutionary time. He termed this evolutionary pattern “sexually dimorphic gigantism.” Gigantism may evolve in Nephilid females in order to increase fecundity, or the number of eggs the female can carry (Lupše et al., 2016). In the Argiopines, while female and male size changes were positively correlated, there were no significant increases in body size. In fact, SSD appeared to decline over time. Dr. Kuntner also wanted to know if the evolution of body size was directional. He tested three evolution models: Brownian motion, directional evolution, and single optimum. In Nephilids, he found that size evolution of females was not directional and fell under the“Brownian motion” model, meaning the changes in size over time were random. Nephilid male size evolution,
Additionally, Dr. Kuntner has done a lot of work on the different morphological and behavioral phenotypes that relate to SSD. He found that complexity in male and female genitalia coevolve (Kuntner, Coddington et al., 2009). Kuntner’s group determined that as female Nephilids evolve to increase fecundity by growing larger and developing ways to mate with multiple males, males respond by developing ways to prevent females from mating with other males. A male’s reproductive fitness increases when he prevents a female from mating with other males by ensuring that she only bears his offspring. One such strategy, called “plugging,” occurs when males essentially castrate themselves and plug the female genitalia to keep other males from being able to mate with her. Other strategies include “mate binding,” which is when a male spins a web around a female to keep her
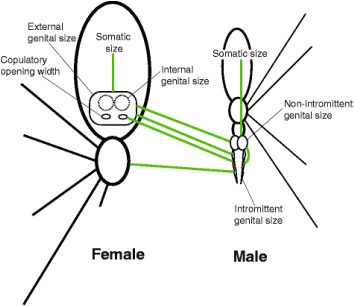
Dr. Kuntner’s more recent work has shifted from studying the evolutionary arms race between male and female orb weavers to looking at the perils of gigantism in female orb weavers. How big is too big? How is mating still possible with females that are so much larger than males? To answer these questions, Kuntner and his team measured hundreds of spider bodies and spider private parts (Lupše et al., 2016). They determined that female body size (or somatic size) evolved independently from internal genital size. Similarly, male body size evolved independently from intromittent (sperm delivering) genital size. However, male intromittent genital size is correlated with female external genital size which indicates that the evolution of these reproductive organs in males helps prevent genital size mismatches that would be expected as a result of SSD (Fig. 3). Dr. Kuntner plans to continue to explore the perils of gigantism in female orb weavers. There are many research questions that the mechanism of SSD poses, such as how gravity affects these large female spiders and how they have had to alter the web designs and functionality to compensate for increases in size. About the Authors: Grace Anderson is a Master’s student in the Shultz Lab at in the UMD Department of Entomology. Her work focuses on the evolution, behavior and systematics of harvestmen. She also is the Assistant Director of the Entomology Department’s Bug Camp. Andrew Garavito is a Master’s student in Dennis vanEngelsdorp’s Lab. He is studying honey bees, with a focus on how pollen consumption impacts the ability of bees to fight off infections. References: Cheng, R.C. & Kuntner, M. 2014. Phylogeny suggests non-directional and isometric evolution of sexual size dimorphism in argiopine spiders. Evolution. doi:10.1111/evo.12504 Hormiga, G., Scharff, N., & Coddington, J. A. (2000). The phylogenetic basis of sexual size dimorphism in orb-weaving spiders (Araneae, Orbiculariae). Systematic Biology, 49(3), 435-462. Kuntner, M., Agnarsson, I. & Daiqin Li. 2015. The eunuch phenomenon: adaptive evolution of genital emasculation in sexually dimorphic spiders. Biological reviews 90: 279–296. Kuntner, M., Arnedo, M.A., Trontelj, P., Lokovšek, T. & Agnarsson, I. 2013. A molecular phylogeny of nephilid spiders: Evolutionary history of a model lineage. Molecular Phylogenetics and Evolution 69: 961–979. Kuntner, M. & Cheng, R.C. 2016. Evolutionary pathways maintaining extreme female-biased sexual size dimorphism: Convergent spider cases defy common patterns. In: Pontarotti, P. (ed) Evolutionary biology: Convergent evolution, evolution of complex traits, concepts and methods. Springer International Publishing, Cham, pp 121–133. Kuntner, M., Coddington, J. A. & Schneider, J. M. 2009. Intersexual arms race? Genital coevolution in nephilid spiders (Araneae, Nephilidae). Evolution 63: 1451-1463. Kuntner, M., Kralj-Fišer, S., Schneider, J. M. & Li, D. 2009. Mate plugging via genital mutilation in nephilid spiders: an evolutionary hypothesis. Journal of Zoology 277: 257-266. Kuntner, M. & Elgar, M. A. 2014. Evolution and maintenance of sexual size dimorphism: Aligning phylogenetic and experimental evidence. Frontiers in Ecology and Evolution 2: 26. Li, D., Oh, J., Kralj-Fišer, S. & Kuntner M. 2012. Remote copulation: male adaptation to female cannibalism. Biology letters 10.1098/rsbl.2011.1202 Lupše, N., Cheng, R.C. & Kuntner, M. 2016. Coevolution of female and male genital components to avoid genital size mismatches in sexually dimorphic spiders. BMC Evolutionary Biology 16:161 Nessler, S. H., Uhl, G., & Schneider, J. M. (2009). Sexual cannibalism facilitates genital damage in Argiope lobata (Araneae: Araneidae). Behavioral Ecology and Sociobiology, 63(3), 355-362. Comments are closed.
|
Categories
All
Archives
June 2024
|


 RSS Feed
RSS Feed



