
|
written by: Kathleen Evans While there remains more to learn about the functional role that biodiversity plays in agroecosystems, it is evident that high-diversity ecosystems promote important ecosystem services such as decomposition and nutrient cycling, pollination, and biotic pest control (Tscharntke et. al. 2005). Biodiversity is thought to provide ecosystem stability and resilience to disturbance, but such evidence is scarce. Significant losses in biodiversity occurred during the post-war era when agriculture practices transformed from traditional to modern and rapid agriculture intensification practices created more homogeneous and simplified landscapes (Tscharntke et. al. 2005; Vandermeer 1997). Unfortunately, there is still a knowledge gap in our understanding of how changes in land-use and simplification have altered the local and regional ecological processes, community structure and ecosystem services. Interaction networks enable us to understand how ecological communities assemble and function and is a useful tool for measuring changes in the stability and resilience of ecological communities. Dr. Adam Vanbergen, Director of Research (écologie des insectes) UMR Agroécologie in Dijon, France, studies interspecific interaction networks (e.g. plant- pollinator) and ecosystem services (e.g. pollination) and how they are affected by environmental change and kindly shared his work with the UMD Entomology Department at our weekly seminar series. Interaction networks describe species patterns and associations and help us to understand the underlying ecological processes behind these interactions. Land-use and other types of human disturbances have consequences for the structure and function of biotic interactions (organisms living in the same community that directly or indirectly influence one another). For example, they can alter pollinator species abundance and diversity, which in turn can affect plant reproduction (e.g plant mating systems) and persistence. Many plant species require a level of outcrossing through the exchange of pollen with another individual of the same species to reproduce and maintain their genetic health. Pollinator behavior, diversity and visitation (number of visits to a flower) can have profound impacts on such mating systems affecting plant reproductive success, ultimately with long-term implications for population persistence of the plant. This hypothesis on the effect of land-use on biotic interactions and plant mating success was tested at a regional scale in Northern Scotland with a project investigating how disturbance of woodland birch habitat patches by the long-term introduction of grazing cattle modified woodland floral resources, plant-pollinator networks and the Cirsium palustre (marsh thistle) mating system (Vanbergen et al 2014). C. palustre (Figure l) was the chosen target species to measure outcrossing rates because it is dependent on insect pollinators for outcrossing and it occurred in both grazed and un-grazed habitat. Ten sites from 3,000 deciduous woodland fragments, five grazed and five un-grazed, were randomly sampled by employing transects and identifying plant and pollinator species, abundance and pollinator visitation frequency to C. palustre and outcrossing levels throughout the season in each transect. With this data, they were able to model the interaction networks of C. palustre. The results showed that grazing sites contained more flowering plant species, thus creating more food resources (pollen and nectar) for pollinators, and in turn creating a larger and more diverse plant-pollinator network (Figure 2). Although the community was more diverse, they found that grazing fundamentally altered the organization of plant-pollinator networks by altering the diversity of the flowering plants. They also found that these network changes altered the C. palustre mating system by increasing the number of flower visitors and thus increasing outcrossing rates (Vanbergen et al 2017). The stability of nested networks was measured using a tool that is referred to as "nestedness". Nestedness is a metric of stability that measures the redundancy of interactions (e.g. plant-pollinator interactions). For example, they measured how many visitors to each of the floral communities in each treatment (grazed and ungrazed sites). They found that the networks in grazed habitats were less nested and the lower nestedness may also suggest that the grazed networks might be vulnerable to extinction cascades. Overall, this study showed how anthropogenic disturbance of habitat can alter the way that communities are organized and ultimately the persistence of wild plant populations. The next step was to test and interpret network structures and stability on a national scale. Using a 30-year dataset of bee, hoverfly and butterfly occurrence records coupled with flower visitation data, Dr John Redhead at the UK Centre for Ecology & Hydrology, supported by Adam and colleagues, was able to model binary plant-pollinator networks and their response to land-use (e.g. agriculture). Using this dataset along with the computer program FRESCALO to estimate species richness, they predicted the structure of plant-pollinator networks on a 10 x 10 km landscape scale in Great Britain (Redhead 2018). With such a large dataset, they could ask questions regarding the spatial variation of plant-pollinator network structure and measure the impacts of human disturbances (agricultural land-use) on the network robustness to extinction across the British national landscape. In summary, they found that network robustness and structure varied spatially (Figure 3) and concluded that, surprisingly, network generality and robustness to extinction increased with percent agricultural land cover in Great Britain. They also found that crop pollinators (managed and wild) were generalist species that are more widely occurring compared to non-crop pollinators, and this subset of the overall network in each landscape was more robust to extinctions. It is important to note that there were significant losses in biodiversity following World War II and consequently, the results of this study reflect a more generalized (generalist) community of pollinators which are able to cope with intensive changes in landscapes compared to more specialized species. It is clear that conventional agricultural intensification was and remains a major driver of species loss, particularly rare species (IPBES 2019). 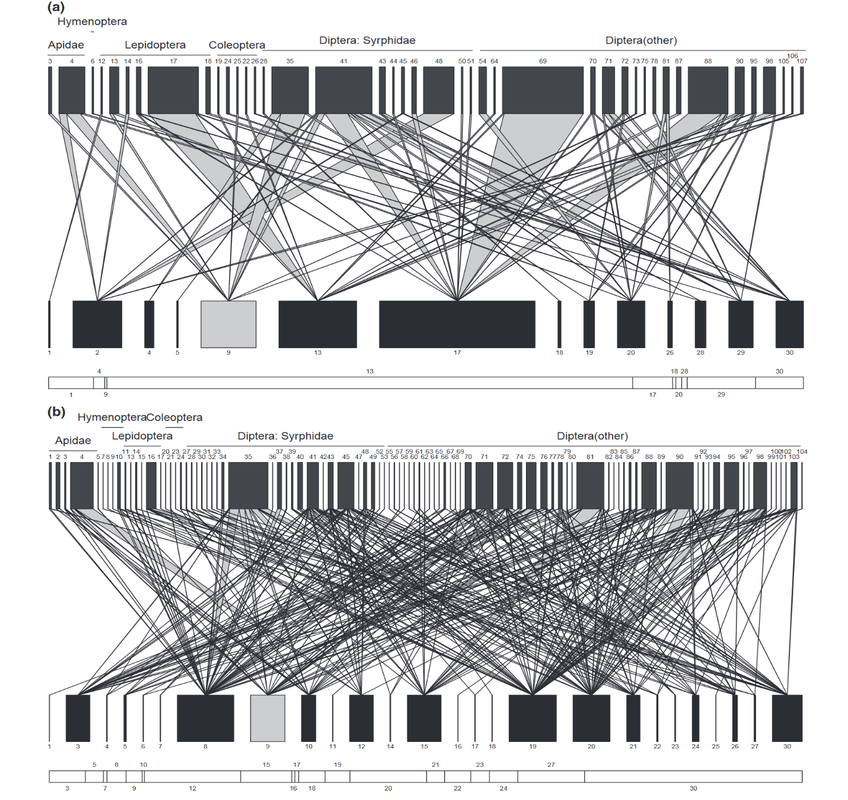 Figure 2. Insect–plant visitation networks summed across (a) four ungrazed and (b) five grazed birch woodlands. The upper and lower levels of the networks represent insects and plants respectively. The thickness of individual bars (i.e. a single species) is proportional to the summed visitation frequency for that species in the network. The lines between the two levels of the network represent the interactions between plant and insect species pairs with line thickness proportional to increasing frequency. (Vanbergen 2014) It is important to note that rare and/or specialized species interactions are often essential to ecosystem functioning by supporting wider biodiversity and that environmental changes (e.g. climate change or habitat loss) and disturbances remove more vulnerable (i.e. rare and less robust) species, causing drastic changes and homogenization in community structure. Another risk comes from future environmental stressors or their combinations (e.g. land-use, climate change, emerging diseases) which may impact these homogenized communities and even eliminate more generalist species. These results and other projects inspired an additional ongoing collaboration between Adam and a group of European Union funded researchers to investigate the viral eco-evolutionary dynamics of wild and domestic pollinators under global change (VOODOO). Specifically, they are investigating plant-pollinator-virus interaction networks in human-dominated landscapes (urban and agricultural). Overall, there is compelling evidence that land-use modifies the structure and function of plant-pollinator networks, raising concerns and questions over how and if ecological processes will adapt to the Anthropocene. 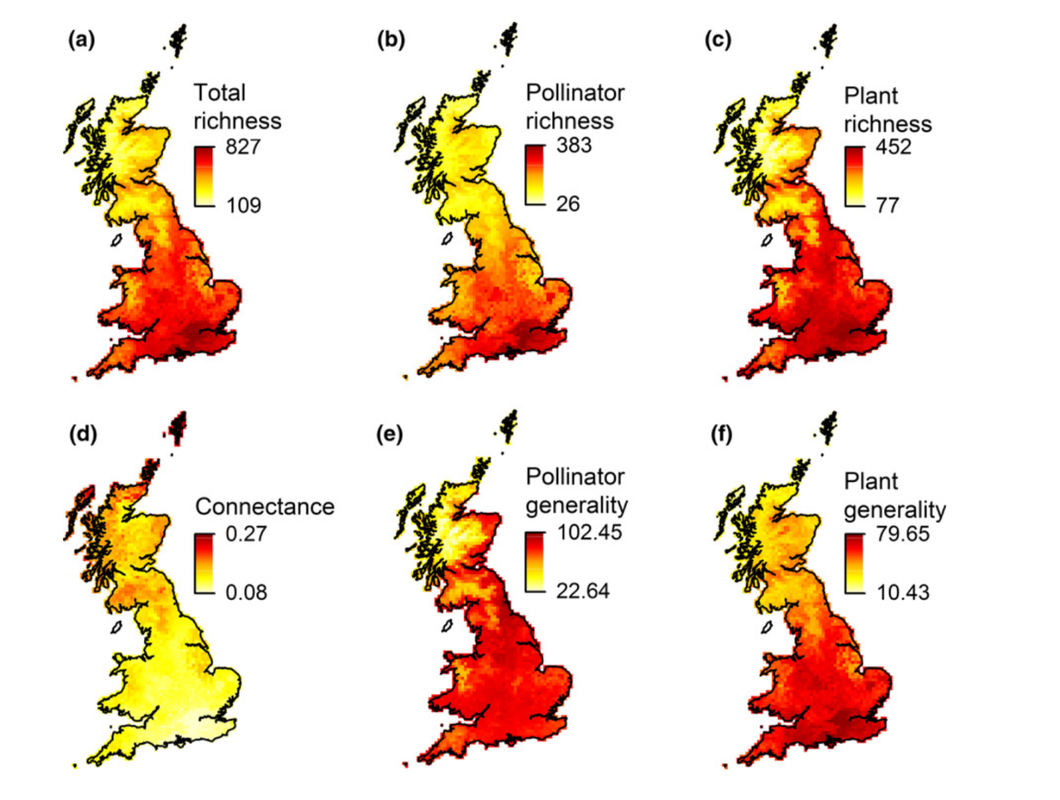 Figure 3. Network properties across GB: a) total species richness of plants and pollinators combined, b) richness of pollinators and c) richness of plants, d) network connectance, e) pollinator generality, and f) plant generality. Lighter colors indicate lower values, darker colors indicate higher (Redhead 2018). Author: Kathleen Evans, PhD student in EspíndoLab studying mutualistic interactions in agroecosystems
References: IPBES. (2016). Assessment Report on Pollinators, Pollination and Food Production. IPBES. http://ipbes.net/assessment-reports/pollinators D., Winfree, R., Bartomeus, I., Carvalheiro, L.G., Henry, M., Isaacs, R. et al. (2015). Delivery of crop pollination services is an insufficient argument for wild pollinator conservation. Nat. Commun., 6, 7414 Redhead, J. W., Woodcock, B. A., Pocock, M. J. O., Pywell, R. F., Vanbergen, A. J., & Oliver, T. H. (2018). Potential landscape‐scale pollinator networks across Great Britain: Structure, stability and influence of agricultural land cover. Ecology Letters, 21(12), 1821–1832. https://doi.org/10.1111/ele.13157 Nentwig W, Müller E (2011) Plant pathogens as biocontrol agents of Cirsium arvense – an overestimated approach? NeoBiota 11: 1-24. https://doi.org/10.3897/neobiota.11.1803 Vanbergen, A. J., Woodcock, B. A., Gray, A., Grant, F., Telford, A., Lambdon, P., Chapman, D. S., Pywell, R. F., Heard, M. S., & Cavers, S. (2014). Grazing alters insect visitation networks and plant mating systems. Functional Ecology, 28(1), 178–189. https://doi.org/10.1111/1365-2435.12191 Comments are closed.
|
Categories
All
Archives
June 2024
|


 RSS Feed
RSS Feed



