
|
Mechanisms regulating condition-dependent growth in the elaborate weapons of sexual selection in beetles  Figure 1. Elk in the Snow (CC 2.0 by Larry Smith) Figure 1. Elk in the Snow (CC 2.0 by Larry Smith) Some of the most charismatic and recognizable features in the animal kingdom are the impressive weapons they wield when competing with each other. Porcupine quills, the chelae of crayfish, and the antlers of elk all require an enormous amount of energy dedicated primarily to fighting their fellows for resources and mates. Most species get by with a modest arsenal; mockingbirds will pick fights over the best territory, but their beaks are hardly fearsome. For centuries scientists have studied and debated the evolutionary pressures that guide the creation of extreme biological weaponry we see in mastodons, kudu, and stalk-eyed flies1. Why do a few species develop these exaggerated armaments when most get by with beaks and jaws that can crack seeds not bones?Some of the most charismatic and recognizable features in the animal kingdom are the impressive weapons they wield when competing with each other. Porcupine quills, the chelae of crayfish, and the antlers of elk all require an enormous amount of energy dedicated primarily to fighting their fellows for resources and mates. Most species get by with a modest arsenal; mockingbirds will pick fights over the best territory, but their beaks are hardly fearsome. For centuries scientists have studied and debated the evolutionary pressures that guide the creation of extreme biological weaponry we see in mastodons, kudu, and stalk-eyed flies1. Why do a few species develop these exaggerated armaments when most get by with beaks and jaws that can crack seeds not bones? It has long been established that the primary driver behind these exaggerated weapons is sexual selection. Sexual selection is the concept that a trait evolves not because it allows the animal to survive in the environment, but because it makes the animal more likely to earn the opportunity to have sex, and therefore offspring. These features are typically used to fight for direct access to mates or ownership of attractive territory, or sometimes just to impress their intended. We consider these traits to be “exaggerated” when they are: disproportionately larger than other traits, cost disproportionately more than other traits, and are more sensitive to environmental conditions than other traits (Condition Sensitivity). Dr. Lavine leads a research group working to identify the chemical and genetic mechanisms which regulate the growth of these structures in insects to better understand what’s driving the evolution of these intriguing structures. Insects are an ideal model organism for studying how these structures develop. Their growth from egg to adult has clearly defined steps which are apparent each time the insect molts. The speed and timing of changes going on inside an insect’s body are relatively regular and easy to determine and manipulate. Furthermore, insects that pupate cannot feed during this process, so they must obtain all of their nutrition as larvae, stop feeding, and then build their entire adult body with this stored energy. This makes them ideal models for studying the effects of nutrition on growth. For their study model Dr. Lavine’s research team selected the stag beetle Cyclommatus metallifer. Males of this species grow enormous mandibles which they use to grapple with other males for access to females. These mandibles develop as the beetle is pupating from the juvenile larva into the adult beetle. 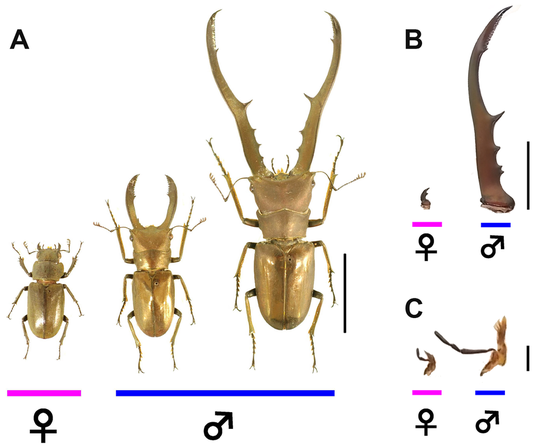 Figure 2. Sexual dimorphism and male variation in C. metallifer. (CC by Gotoh, 2014) These impressive structures are ideal examples of exaggerated sexually dimorphic weaponry. As you can see in Figure 2, the body of the male on the right is only slightly larger than the left male, but his mandibles are disproportionately larger. Previous work had indicated that a chemical known as Juvenile Hormone (JH) can play a role in the disproportionate development of various features in ants, bees2, and other beetles3. To investigate JH’s role in regulating mandible growth in C. metallifer Dr. Lavine’s group exposed developing male beetles to a synthetic chemical called JH-analog (JHA). They found that exposing males to high levels of JHA during the larval stage increases the entire body size, while exposure slightly later, at the early prepupal stage, increased mandible size without regard to body size4. This shows that beetle mandibles are very sensitive to JH signaling during a specific time in their development and that not all traits respond in the same way to JH. But what causes the level of JH to differ between beetles? JH levels are linked to nutrition, and indeed when Dr. Lavine’s group raised beetle larvae with small amounts of food the level of JH in their hemolymph during that critical early prepupal period was lower than that of larvae raised with plenty of food4. Male and female beetles have similar levels of JH during development. If JH levels explain the differences between large and small males, what is driving the difference between males and females? The logical explanation is that male mandible tissue has a unique sensitivity to JH. To determine why female mandibles aren’t sensitive to JH Dr. Levine’s group decided to look the relationship between JH and the C. metallifer doublesex gene (Cmdsx), which has been shown in other organisms to play a role in the physiological differences between the sexes5. Gene “knockdown” experiments allow you to characterize an organism’s phenotype when the expression of the target gene is absent. By comparing organisms with a functioning gene to individuals where the expression is absent we can understand its function. To do this, Dr. Lavine’s group used a technique known as RNA-interference (RNAi) to knockdown the Cmdsx gene in male and female beetles6. They found that males without functioning Cmdsx genes didn’t grow impressive mandibles, and females without working Cmdsx genes when mature were the golden metallic color of a male beetle. These along with assorted other features clearly showed that interfering with the function of Cmdsx genes produces individuals which are not characteristic of their sex, or “intersex.” The next step was to illustrate the link between Cmdsx and JH by applying JH to these intersex beetles during that critical prepupal period. The females without functioning female Cmdsx suddenly became sensitive to JH, growing larger mandibles, while the males lost most of their sensitivity. They conclude that the default state of a beetle mandible is moderate JH sensitivity, with female Cmdsx inhibiting sensitivity, and male Cmdsx enhancing it. This was the first study to provide a causal link between secondary-sexual characteristics and nutritionally dependent JH signaling pathways. Dr. Lavine’s work on the genetic and physiological mechanisms underlying sexually dimorphic beetle weapons has important impacts on understanding how these sorts of traits evolve across the animal kingdom. The JH/Cmdsx system is one of only a small handful that have been identified to control the growth of these exaggerated attributes, but it is already becoming clear that each mechanism is coupled with specific functions. The pathways that have been used to amplify sexually selected traits function very differently than those regulating other extreme traits that aid in hunting or movement7. Understanding how these mechanisms function expands our understanding of the roles of these extreme features and how they evolve. About the Authors Peter Coffey is a Master's student whose research focuses on using cover crops to optimize sustainable farming economics. His current projects using lima beans and eggplant as model systems focus on plant nutrition, weed suppression, influences on pest and beneficial insects, and crop yield value. Follow him on twitter at @petercoffey. Mengyao Chen is a Master’s student in Dr. Leslie Pick’s Lab. Her research focuses on segmentation genes in Brown Marmorated Stink Bug (BMSB, Halyomorpha halys). Her current work is looking for orthologs of pair-rule genes in BMSB, and studying their expression and functions using RNA in situ hybridization and RNAi. The authors would like to acknowledge Drs. Kelly Hamby and Laura Lavine for their comments and feedback on this piece. References:
1. Lorenz, K. On Aggression. 16th Ed. New York: Bantam Books; 1966 2. Hartfelder K, Emlen DJ (2005) Endocrine control of insect polyphenism. In: Gilbert LI, Iatrou K, Gill SS, editors. Comprehensive Molecular Insect Science. Oxford: Elsevier-Pergamon. pp. 651–703. Available:dbs.umt.edu/emlenlab/documents/Emlen%20Publications/hartfelderEmlen.pdf 3. Emlen DJ, Allen CE (2003) Genotype to phenotype: physiological control of trait size and scaling in insects. Integ Comp Biol 43: 617–634. 4. Gotoh H, Cornette R, Koshikawa S, Okada Y, Lavine LC, et al. (2011) Juvenile hormone regulates extreme mandible growth in male stag beetles. PLoS ONE 6(6): e21139 Available: http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0021139. 5. Tanaka K, Barmina O, Sanders LE, Arbeitman MN, Kopp A (2011) Evolution of sex-specific traits through changes in HOX-Dependent doublesex expression. PLoS Biol 9(8): e1001131 Available: http://www.plosbiology.org/article/info%3Adoi%2F10.1371%2Fjournal.pbio.1001131. 6. Gotoh H, Miyakawa H, Ishikawa A, Ishikawa Y, Sugime Y, et al. (2014) Developmental Link between Sex and Nutrition; doublesex Regulates Sex-Specific Mandible Growth via Juvenile Hormone Signaling in Stag Beetles. PLoS Genet 10(1): e1004098. doi: 10.1371/journal.pgen.1004098 Available: http://www.plosgenetics.org/article/info%3Adoi%2F10.1 371%2Fjournal.pgen.1004098 7. Laura Lavine, Hiroki Gotoh, Colin S. Brent, Ian Dworkin, and Douglas J. Emlen. (2015) Exaggerated Trait Growth in Insects. Annual Review of Entomology Vol. 60: 453-472 Available: http://www.annualreviews.org/doi/abs/10.1146/annurev-ento-010814-021045 Additional Reading: 1. Emlen DJ, Warren I, Johns A, Dworkin I, Corley Lavine L. 2012. A mechanism of extreme growth and reliable signaling in sexually selected ornaments and weapons. Science 337: 860-864. DOI:10.1126/science.1224286. 2. Video: http://video.sciencemag.org/VideoLab/beetles/ 3. Emlen DJ. Animal Weapons: The Evolution of Battle. New York: Henry Holt and Co; 2014. Available: www.amazon.com/Animal-Weapons-Evolution-Douglas-Emlen/dp/0805094504/ref=tmm_ hrd_swatch_0?_encoding=UTF8&sr=&qid= Comments are closed.
|
Categories
All
Archives
June 2024
|
 RSS Feed
RSS Feed



