
|
Many people are fascinated about the diversity in nature, some are attracted by cichlids with various colors, others were allured by butterflies with different patterns. Katherine Taylor, a new Post-doc in the Fritz lab, was interested in the diversity of the lacewing mating songs. Lacewings create their mating songs by vibrating their abdomens. These intermittent vibrations propagate through the substrate they are on and are received by the possible mates. The mating “song” is not a single burst of vibrations; there is a certain amount of time between each song burst which can be referred to as the volley period. The vibration is transmitted through their legs to the leaf, then detected by potential mates. The potential mates sing nearly identical songs that are performed together in a duet prior to mating. Interestingly, this vibrational signal differs among lacewing species, and this is what Dr. Taylor was fascinated about during her Ph.D. years at the University of Connecticut. Lacewings are major generalist predators, which are able to prey on multiple pest species including aphids, scale insects, moth eggs, etc [1][2] (Figure 1). Lacewings have a wide Holarctic distribution. It was once thought that many green lacewings belonged to a single species until the diversity of their mating songs was discovered by Dr. Charles Henry, Dr. Taylor’s dissertation advisor.  Figure 1: A green lacewing larvae is preying on aphids. In order to identify the genetic basis of lacewing mating songs, Dr. Taylor performed an experiment with lacewings from different species: one species produces a volley period which is twice as long as the other. She performed a hybrid cross of those two lacewing species and recorded the volley periods of both parents and the first-generation offspring (F1). The second generation (F2) was produced by crossing within F1, with their volley periods were also recorded. She found the F1 had an intermediate volley period and F2 retained this trait, but with a slightly wider range compared to F1 (Figure 2). Dr. Taylor also did an experiment to identify the relationship between sound production and preference in of the F2 lacewings. She found a correlation between the signal produced and preferred in both male and female lacewings in the F2 generation, indicating that the mating signal and preference in these lacewings are genetically linked. In order to identify genes that contribute to lacewing songs, she then sequenced the genome of 80 F2 individuals, as well as two parents using restriction site-associated DNA sequencing (RAD sequencing). She found one region in the lacewing genome which might be associated with the volley period. She also performed a divergence scan, a technique to test if a gene locus is under selection, and she found that the same region was under selection. Later, she performed a genome assembly and annotation, and identified 348 putative genes in the region, with three related to mating behavior, and one related to wing generated song production[1]. 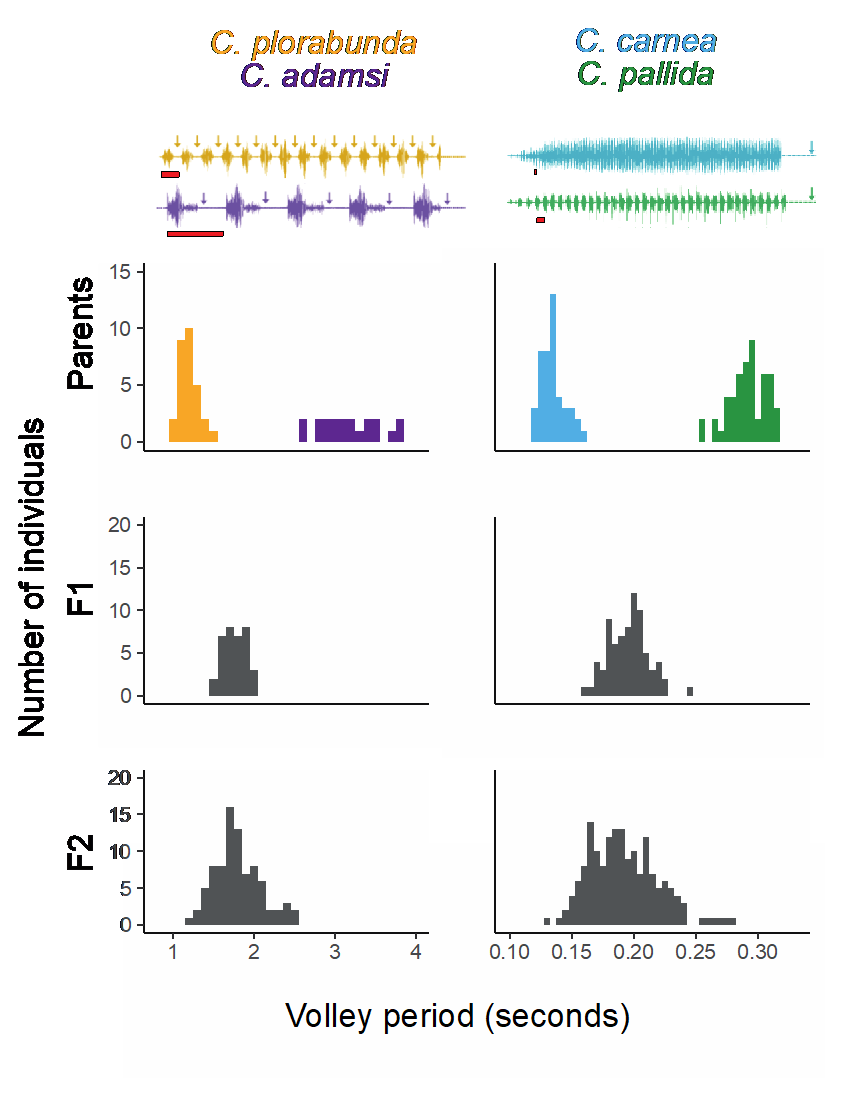 Figure 2: The volley periods of two lacewing species, their first-generation offspring (F1) and the second generation (F2). Figure 2: The volley periods of two lacewing species, their first-generation offspring (F1) and the second generation (F2). While studying lacewings, Dr. Taylor noticed lacewing larvae are able to perform death feigning. Death feigning is defense mechanism of organisms to playing dead, and it is a common behavior in many animals, for instance, beetles, possums, etc. Dr. Taylor decided to test what might be the cause of lacewings’ death feigning. To test if it has anything to do with the energetic sources available to the insect, she separated lacewing larvae into three groups: one group had access to food at all times while the other two groups were starved for one and two days. She used fingers to poke each larvae up to twenty times and recorded how many stimulation times it will take for each larva to do death feigning. Interestingly, they found out the individuals from the group which was not exposed to any starvation required more stimulation before death feigning compared to the other two groups. As a result, she concluded that death feigning is used when larvae lack sufficient energetic resources to engage in more costly defensive strategies such as running away or fighting back. To test if this variation in an individual’s death feigning ability is merely caused by environmental conditions, she decided to check if there are any underlying genetic factors. She crossed lacewings which were more likely to death feign with lacewings that are less likely to death feign and checked if their offspring used death feigning as a defense strategy. She found a mean heritability of 0.53 in offspring, which indicates this defense strategy is heritable thus controlled by genetic factors.
In the future, Dr. Taylor will collaborate with Dr. Megan Fritz to identify possible genes associated with Bt crystal toxin resistance in Helicoverpa zea, the corn earworm. Corn earworms have been reported to have developed Bt resistance in recent years. Dr. Taylor is going to identify additional resistance-associated genomic regions using RAD sequencing to then identify possible genes that contribute to this Bt resistance in corn earworms. Authors: Huiyu Sheng is a third-year master student in the St. Leger lab studying Metarhizium endophytic ability. Serhat Solmaz is a second year MS student in the vanEngelsdorp Lab studying the pathophysiological effects of Nosema ceranae on honey bee workers at the University of Maryland Reference: [1] K. Taylor, “Speciation Genomics of the Chrysoperla carnea Complex,” Dr. Diss., May 2020. [2] “Green lacewing larvae vs. bird cherry-oat aphid - YouTube.” [Online]. Available: https://www.youtube.com/watch?v=7jFAkOX8y3s. [Accessed: 24-Sep-2020]. Comments are closed.
|
Categories
All
Archives
June 2024
|
 RSS Feed
RSS Feed



